The order Condylarthra is one of the most characteristic groups of Paleocene mammals, and it illustrates well the evolutionary level of the Paleocene mammal fauna. When compared to the mammal fauna of today, condylarths are relatively unspecialized placental mammals. However, in comparison to their insectivorous ancestors, members of the Condylarthra show the first signs of being omnivores or even herbivores. Since larger herbivores were absent on land after the extinction of the dinosaurs, this shift in diet triggered the tremendous evolutionary radiation of the condylarths that we can observe throughout the Paleocene. Result of this radiation are the different groups of ungulates (or "hoofed mammals") that form the dominant large herbivores in most Cenozoic animal communities on land, except on the island continent of Australia. The term ungulate refers here to a subgroup of placental mammals (the Ungulata) that are descendants of a common ancestor, the most primitive condylarth. Among recent mammals, the even- and odd-toed ungulates, hyraxes, elephants, aardvarks, sea cows and whales are traditionally regarded as members of the Ungulata (but see discussion at the end of this article). Besides condylarths several extinct groups must be added to the Ungulata, especially the endemic South American orders of ungulates. Although many ungulates have hoofs, this feature does not define the Ungulata. Some condylarths indeed have small hoofs on their feet, but the most primitive forms are clawed. On the other hand, hoofs have been independently acquired by groups that do not belong to the Ungulata, such as the extinct Pantodonta.
The majority of condylarths is known from North America, the continent that has the most complete record of Paleocene mammals. Only few Paleocene mammal faunas have been discovered in Europe, but these show that condylarths were equally important in this part of the world and were often represented there by close relatives of North American forms. Surprisingly, only a few questionable condylarths have been described from the much richer Paleocene faunas of Asia. The ecological role of plant-eating mammals was taken over there by groups like the Anagalida and the Pantodonta. Yet these were hunted by carnivorous descendants of the condylarths, members of the order Mesonychia. South America is the only continent in the southern hemisphere that has a substantial record of Paleocene mammals, and the growing number of South American condylarths establishes an important link to the northern faunas.
The most primitive known condylarth is the rat-sized Protungulatum ("before-ungulate") from the United States and Canada. Besides its occurrence in clearly early Paleocene sediments, remains of this genus are also found together with the teeth of dinosaurs. Therefore it was originally thought that Protungulatum (and some other types of mammals) appeared in the latest Cretaceous and co-existed with the last dinosaurs and with typically Cretaceous mammals in the so-called "Bugcreekian" faunas. In this scenario, early ungulates like Protungulatum would have been in competition with the herbivorous dinosaurs and could have contributed to the demise of these reptiles. However, a more recent interpretation is that the "Bugcreekian" assemblages are a mixture of early Paleocene and latest Cretaceous fossils, the latter having been eroded by rivers from older sediments and redeposited together with fresh animal remains in the early Paleocene. The issue is not finally resolved, but this new interpretation fits to the idea that mammals filled the vacant ecological niches after a catastrophic extinction of the dinosaurs, which may have been caused by the impact of a large meteorite. Whatever the precise age of the fossils, the status of Protungulatum as prototype for the later ungulates has not been challenged since its first description in 1965. Although still quite primitive, the dentition of Protungulatum foreshadows the typical trends observed in ungulates: Its teeth are low and have increased potential for crushing and grinding food, which probably allowed the animal to feed on soft plants, fruits and still some insects. Only few bones are known for these earliest condylarths beyond their jaws and dentition, so a lot remains to be learnt about their biology. Yet it is a big progress that potential ancestors of Protungulatum have recently been identified in the late Cretaceous of Uzbekistan. These small placental animals, called zhelestids, are not strictly classified as ungulates, but their teeth may show us a first stage in the evolution of an ungulate-like dentition.
Protungulatum is traditionally regarded as a member of the Arctocyonidae, a family that was diverse and abundant in the Paleocene of North America and Europe. The arctocyonids are the least herbivorous group of condylarths. In fact their skulls look superficially like those of carnivores, with large canines and relatively sharp teeth, but arctocyonids had no specialized teeth for slicing meat and were probably omnivores. The limbs of arctocyonids were relatively short and showed none of the specializations that we typically associate with ungulates, like reduction of the side digits, fusion of bones or the possession of hoofs. Many arctocyonids are only known from their teeth, which show much individual variation, so the taxonomy of arctocyonid genera and species is highly unstable.
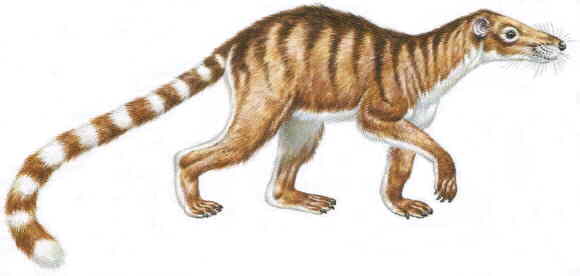
Figure 1: Reconstruction of the agile climber Chriacus, a small arctocyonid from the early Paleocene to early Eocene of North America. Length including tail about 1m. From Cox (1988). See also reconstruction in a middle Paleocene forest.
With an estimated body mass of 5 to 10 kg, Chriacus from the early Paleocene to early Eocene of North America is an example of the smaller arctocyonids. A nearly complete skeleton has been found in early Eocene rocks of Wyoming. It is essentially primitive in structure and similar to early Paleocene arctocyonids like Loxolophus that are less well known. As this skeleton shows, Chriacus was equally adept in the trees and on the ground. Among todays mammals it can be compared best to members of the racoon family and to the civets. Like most climbing mammals, Chriacus had powerful limb musculature, very mobile joints and feet bearing five digits with claws. The tail of Chriacus was long and robust. It was well adapted to be used in balancing and may even have formed a prehensile organ. As the design of the forelimb suggests, the animal was capable of digging. Chriacus may have eaten fruits, insects and other small animals. Interestingly, its lower incisors formed a kind of tooth-comb that was probably used for grooming.
Although we can presume that larger arctocyonids like Arctocyon and Claenodon were still able to climb in trees, they obviously spent most of their time on the forest floor. Ecologically these animals may have been ecologically comparable to the bears. They may have partly taken over the role of large predators in Paleocene faunas, together with two other families of ungulates that evolved from early arctocyonids, the triisodontids and mesonychids. The latter two groups were traditionally also classified as condylarths but are now regarded as members of the order Mesonychia. All these carnivore-like ungulates are treated in a separate article on early predatory mammals.
Besides the arctocyonids, members of the Periptychidae were the dominant condylarths of early Paleocene faunas in North America (the so-called Puercan fauna). Less than one million years after the end of the Cretaceous they already span a wide range of size, from squirrel-sized forms like Anisonchus to the sheep-sized Ectoconus, one of the largest Puercan mammals. A complete skeleton of Ectoconus ditrigonus has been found in New Mexico, U. S. A., making this species the best known mammal of the Puercan fauna. Incidentally the skeleton shows a severely diseased elbow joint, which must have made the animal lame. The general body plan of Ectoconus agrees in many points with the larger arctocyonids, like in the massive build, the small braincase of the skull, the stout short limbs and the long heavy tail. However, the five wide-spreading digits on its feet carried small hoofs similar to those of the recent tapirs, in contrast to the clawed feet of the arctocyonids. Although Ectoconus is a nearly ideal approximation of the generalized ungulate body plan, the dentition of the periptychids is specialized in a peculiar way, which disqualifies them from being potential ancestors of the modern ungulates. Most periptychids have enlarged, bulbous premolars that probably served for shredding tough plant material. Strong grooves run down from the top of these teeth in the last surviving genus, the early to late Paleocene Periptychus, an animal a little smaller than Ectoconus but with a larger head. The characteristic premolars of Periptychus are strikingly similar to those of some pigs, and in fact a pig- or peccary-like diet, mainly vegetarian, has been proposed for the periptychids.

Figure 2: Reconstruction of the sheep-sized periptychid Ectoconus from the early Paleocene of New Mexico, U. S. A. From Williamson & Lucas (1993).
A third family of condylarths, the Hyopsodontidae, got important later in the Paleocene when the periptychids had already passed the zenith of their evolution. Hyopsodontids were typically small animals that had little in common with our general idea of an ungulate and were more like insectivores in appearance. In some cases the resemblance to insectivores even extends to details of the teeth, and there is much debate whether some putative hyopsodontids like Litomylus are actually insectivores related to the hedgehogs. The best known hyopsodontid is Hyopsodus, a highly successful genus that occurs in the early Eocene all over the northern hemisphere and survived far into the Eocene as one of the last condylarths - although this is of course a question of definition of that group. Rare fossils from North America could indicate that Hyopsodus first appeared there in the latest Paleocene. Hyopsodus was a rat-sized gracile animal with shortened limbs and clawed feet. Its skeleton suggests that it was partly living in trees. A European subfamily of hyopsodontids is centered around Louisina from the later Paleocene of Germany and France. The related genus Paschatherium must have been extremely numerous in the latest Paleocene and earliest Eocene of Europe since it makes up the majority of all mammal fossils in some fossil sites. Ankle bones of this form suggest that Paschatherium, too, was versatile in the trees and may have been similar to squirrels in adaptation.

Figure 3: Reconstruction of Hyopsodus, a rat-sized animal from the early to middle Eocene and possibly latest Paleocene. Hyopsodus is the only hyopsodontid for which the skeleton is adequately known. From Savage & Long (1986).
Mioclaenus and allied genera from the Paleocene of North America have long be considered as another hyopsodontid subfamily. Today they are regarded as a separate family of condylarths, the Mioclaenidae. Mioclaenids got slightly larger than the hyopsodontids, up to hare-size. Like in the Periptychidae, enlarged, bulbous premolars are typical of the mioclaenid dentition. They were probably helpful in processing tougher, more fibrous plants. The middle Paleocene genus Mioclaenus carries the enlargement of the premolars to the extreme. In 1888 the famous American paleontologist Edward Drinker Cope placed an impressive number of twenty-six different species in Mioclaenus. Twenty-three of these have since been removed from the genus, which makes Mioclaenus one of the most abused genus names in the history of mammalian paleontology. The remainder of Cope's species are today all regarded as belonging to the type species, Mioclaenus turgidus.
Although no typical mioclaenids are known from Europe, Pleuraspidotherium from the late Paleocene of France may be an aberrant member of this family. Pleuraspidotherium, an animal about 60 cm in head and body length, is one of the most abundant mammals in river deposits near Cernay, which have even been called the Pleuraspidotherium beds. Large herds of this condylarth must have browsed on the vegetation at the riverside. The cheek teeth of Pleuraspidotherium were surprisingly advanced for a Paleocene mammal: They have a so-called selenodont pattern of crescent-shaped ridges, similar to that developed much later by even-toed ungulates like deer and camel. Equipped with such a dentition, Pleuraspidotherium must have been one of the most exclusive herbivores of its time. Concerning its locomotion, however, the animal was still a generalist. Its skeleton is similar to that of arctocyonids like Chriacus but lacks the tree-climbing adaptations of the latter.
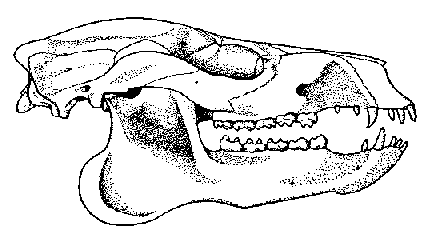
Figure 4: Skull of Pleuraspidotherium aumonieri, an advanced herbivore that is abundant in the late Paleocene of Cernay, France. Length about 13cm. From Russell (1964).
Fossils of mioclaenids have long been restricted to the northern hemisphere, but exciting discoveries have recently shown that these condylarths colonized South America and had their greatest success on that continent. Up to now, South American mioclaenids have only been identified in faunas from the earlier Paleocene, like Molinodus and Tiuclaenus from the important locality of Tiupampa in Bolivia. However, an endemic South American family of condylarths, the Didolodontidae, has been known since a long time from later faunas. Didolodontids and mioclaenids are very similar in their dentition, which suggests that the didolodontids evolved from mioclaenids in South America. Unfortunately, didolodontids are only represented by scanty fossil remains such as teeth, jaws and isolated ankle bones. They were mainly animals of small size, such as Ernestokokenia from the late Paleocene to early Eocene of Argentina, but also included larger forms like the middle Paleocene Lamegoia from Brazil. In their dentition the didolodontids are almost indistinguishable from the most primitive members of the Litopterna, an important order of endemic South American ungulates that later produced horse-like and camel-like forms. In comparison to the didolodontids, the major innovation in primitive litopterns is an advanced structure of the limbs, especially of the ankle. These changes are probably linked to a leaping type of locomotion in the small early litopterns; they allowed later litopterns to become adapted for fast running. Evidence from the dentition suggests strongly that the litopterns, like the didolodontids, have mioclaenid ancestors. To formalize these relationships, it has been recently proposed to define a new order of mammals, the Panameriungulata, for mioclaenids, didolodontids and litopterns. This would at least create a safe home for primitive forms with unknown ankle structure that cannot be confidently assigned to either didolodontid condylarths or litopterns, such as Asmithwoodwardia, a minute animal known from the middle Paleocene of Brazil and the early Eocene of Argentina.
Back in the northern hemisphere, another family of condylarths, the Phenacodontidae, may include the ancestors of a more familiar ungulate order: The odd-toed ungulates or Perissodactyla, represented by horses, rhinos and tapirs in the recent fauna. Historically, phenacodontids form the core of the Condylarthra. Well-preserved skeletons are known for the type genus Phenacodus, which is a good model of an ancestral ungulate with beginning adaptations for running. Unlike arctocyonids, periptychids or mioclaenids, the phenacodontids are not part of the first wave of condylarths that populated North America. They first appear with the fox-sized Tetraclaenodon in the middle Paleocene of that continent. The appearance of the more advanced phenacodontids Phenacodus and Ectocion marks the beginning of late Paleocene time in North America. The type genus Phenacodus covers the large size range of phenacodontids and includes roughly sheep-sized animals. Members of the genus Ectocion were usually smaller, with a body mass of only 3 kg in the smallest species, but there is some overlap in size between the two genera. Phenacodontids were the dominant mammals in the latest Paleocene of North America and account for up to 50% of all mammal specimens in faunas of that age. Both Phenacodus and Ectocion survive until the middle Eocene, but phenacodontids become less common after the end of the Paleocene. Remarkable exceptions are local mass occurrences of the dog-sized phenacodontid Meniscotherium which forms real bonebeds in some places. Meniscotherium is mainly early Eocene in age, although first individuals may already have been present in the latest Paleocene. Phenacodus spread into Europe in the early Eocene as part of a major faunal exchange between the Old and New World, but it never became an important component of the European fauna.

Figure 5: Reconstruction of the late Paleocene to middle Eocene Phenacodus, a sheep-sized herbivore with improved capabilities for running. From Savage & Long (1986).
The skeleton of phenacodontids is generally primitive, especially in the long, heavy tail, but some similarities to perissodactyls are evident in Phenacodus: Its limbs are longer than in primitive condylarths and have five hoofed digits, the first and fifth digit being reduced in size. This foreshadows the early Eocene horse Hyracotherium (sometimes called "Eohippus"), which has completely lost the first digit of the hand and the first and fifth digit of the foot. The limb design indicates that Phenacodus was adapted to running to some degree. Only few limb remains are known for Ectocion, but they suggest that these smaller animals may even have been somewhat better runners than its larger relatives. The skull of phenacodontids is long and has a small braincase. In Phenacodus intermedius the nasal bones are retracted, like in recent tapirs, which may indicate that this species of Phenacodus had a short trunk. As the dentition shows, at least the later phenacodontids were herbivores: Their cheek teeth have low cusps that sometimes tend to become joined into crests, similar to early perissodactyls like Hyracotherium. In Meniscotherium these crests become crescent-shaped, a precocious adaptation for an early Tertiary mammal that can only be compared to the selenodont dentition of Pleuraspidotherium. Although regarded as closely related in the past, later studies have concluded that no special relationship exists between Pleuraspidotherium and Meniscotherium and that the latter is in fact a highly specialized phenacodontid.
As the preceding survey shows, the order Condylarthra contains forms that were adapted to a wide diversity of different ways of life, ranging from squirrel-like tree-dwellers to sheep-sized herbivores and large bear-like omnivores. Although all condylarths share a common ancestor somewhere near Protungulatum, they are only united by their primitiveness in comparison to the more advanced ungulates. For this reason the order Condylarthra has been criticized as a "wastebasket" for primitive forms of ungulates, and it has been proposed to abandon altogether the concept this group. Several alternative orders have been proposed to harbour members of the condylarthra, most recently the Panameriungulata for the branch that led to the South American litopterns, but no consensus about a new systematic arrangement has emerged up to now.
On the other hand, we have to admit that the role of condylarths as universal ancestors of the more modern ungulates is not yet sufficiently documented by the available fossils. In many cases the search for the "missing links" is still ongoing. The Litopterna may be the group of advanced ungulates that is most convincingly rooted in a specific group of condylarths, the mioclaenids. The case of the similarly adapted odd-toes ungulates or Perissodactyla of the northern hemisphere is less clear. If they did indeed evolve from phenacodontid condylarths, perissodactyls must have branched off from a primitive form like Tetraclaenodon, but transitional forms have not yet been found. In addition, an enigmatic skull with strikingly perissodactyl-like teeth has recently been described from the late Paleocene of China under the name Radinskya. Although no limb remains are known for Radinskya, this animal presents a plausible alternative for the origin of the odd-toed ungulates. The relationships of Radinskya to other Paleocene mammals are not clear, but it could belong to an endemic Asian family of herbivores called Phenacolophidae - also of uncertain affinities.
In other cases molecular studies have questioned traditional views. Molecular biologists have recently removed elephants, hyraxes, aardvarks and sea cows from the Ungulata to a new group Afrotheria, an African radiation of placental mammals that also includes tencres, golden moles and elephant shrews (finally united with the name-giving pachyderms). If this is confirmed by future research, living ungulates, and thus still existing descendants of the Condylarthra, would be limited to even- and odd-toed ungulates and whales.
Nevertheless, the recent study of a partial condylarth skeleton shows how a single fortunate discovery can revive old ideas that had already been abandoned. Judging from the teeth, the specimen from the middle Paleocene of New Mexico represents a small arctocyonid close to Chriacus. However, instead of the tree-climbing adaptations known from skeletons of Chriacus, limb fragments of the new specimen indicate initial adaptations for running, strikingly similar to early members of the even-toed ungulates or Artiodactyla. This provides new support for the hypothesis, originally based on dental evidence, that artiodactyls are the descendants of arctocyonid condylarths. At the same time, this surprising discovery demonstrates that Paleocene mammals were much more diverse in adaptations than their jaws and teeth can tell us.
Major sources for this article:
Archibald, J. D. 1996: Fossil Evidence for a Late Cretaceous Origin of "Hoofed" Mammals. Science 272, 1150-1153.
Archibald, J. D. 1998: Archaic ungulates ("Condylarthra"). In: Janis, Ch. M., Scott, K. M. & Jacobs, L. L. (eds.): Evolution of Tertiary Mammals of North America. Volume 1: Terrestrial Carnivores, Ungulates, and Ungulatelike Mammals. Cambridge University Press, 292 - 331.
Carroll, R. L. 1988: Vertebrate paleontology and evolution. Freeman and Company.
Cifelli, R.L. 1983: The Origin and Affinities of the South American Condylarthra and Early Tertiary Litopterna (Mammalia). American Museum Novitates 2722, 1-49.
Cox, B. (editor) 1988: Illustrated Encyclopedia of Dinosaurs and Prehistoric Animals. Macmillan London Limited.
Godinot, M., Smith. T. & Smith, R. 1996: Mode de vie et affinités de Paschatherium (Condylarthra, Hyopsodontidae) d'après ses os du tarse. Palaeovertebrata 25 (2-4), 225-242.
Madsen, O. et al. 2001: Parallel adaptive radiations in two major clades of placental mammals. Nature 409, 610-614.
McKenna, M. C., Minchen, C., Ting, S.Y. & Zhexi, L. 1989: Radinskya yupingae, a perissodactyl-like mammal from the late Paleocene of China. In: Prothero, D. R. & Schoch, R. M. (eds.): The Evolution of Perissodactyls. Oxford Univ. Press, New York, 24-36.
de Muizon, C. & Cifelli, R. L. 2000: The "condylarths" (archaic Ungulata, Mammalia) from the early Palaeocene of Tiupampa (Bolivia); implications on the origin of the South American ungulates. Geodiversitas 22 (1), 47-150.
Murphy, W. J. et al. 2001: Molecular phylogenetics and the origins of placental mammals. Nature 409, 614-618.
Kurtén, B. 1971: The age of mammals. Columbia University Press, New York.
Lofgren, D. L. 1995: The Bug Creek Problem and the Cretaceous-Tertiary Transition at McGuire Creek, Montana. Univ. of California publications in geological sciences 140, 1-185.
Matthew, W. D. 1937: Paleocene faunas of the San Juan Basin, New Mexico. Transactions of the Philosophical Society 30, 1-510.
Rose, K. D. 1981: The Clarkforkian land-mammal age and mammalian faunal composition across the Paleocene-Eocene boundary. University of Michigan, Papers on Paleontology, v. 26, 1-197.
Rose, K. D. 1987: Climbing Adaptations in the Early Eocene Mammal Chriacus and the Origin of Artiodactyla. Science 236, 314-316.
Rose, K. D. 1996: On the origin of the Artiodactyla. Proc. Natl. Ac. Sci. USA 93, 1705-1709.
Russell, D. E. 1964: Les mammifères paléocènes d'Europe. Mémoires du Muséum National d'Histoire Naturelle, Série C, 8, 1-324.
Savage, R. J. G. & Long, M. R. 1986: Mammal evolution. An illustrated guide. British Museum (Natural History).
Thewissen, J. G. M. 1990: Evolution of Paleocene and Eocene Phenacodontidae (Mammalia, Condylarthra). University of Michigan Papers on Paleontology 29, 1-107.
Thewissen, J. G. M. 1991: Limb osteology and function of the primitive Paleocene ungulate Pleuraspidotherium with notes on Tricuspiodon and Dissacus (Mammalia). Geobios 24 (4), 483-495.
Van Valen, L. 1978: The beginning of the Age of Mammals. Evolutionary Theory 4, 45-80.
Williamson, T. E. & Lucas, S. G. 1992: Meniscotherium (Mammalia, "Condylarthra") from the Paleocene-Eocene of western North America. Bulletin of the New Mexico Museum of Natural History and Science 1, 1-75.
Williamson, T. E. & Lucas, S. G. 1993: Paleocene vertebrate paleontology of the San Juan Basin, New Mexico. Bulletin of the New Mexico Museum of Natural History and Science 2, 105-135.