
It is in the early Paleocene that we first encounter animals in the fossil record which show strong links to our own order, the primates. Surprisingly, early primate-like forms are best known from continents that are today, apart from their southernmost corners, not inhabited by primates other than human beings: Europe and North America. This unusual geographical occurrence can be explained by the warm, even subtropical conditions that existed far into the north during Paleocene time. Favorable conditions also allowed other exotic animals like crocodiles to thrive at high latitudes. Yet the importance and diversity of primate-like mammals in the Paleocene faunas of Europe and North America is remarkable.
During the Paleocene most primate-like animals belonged to a group called Plesiadapiformes. Traditionally, the plesiadapiforms have been regarded as archaic members of the order Primates. Although plesiadapiforms are similar to modern primates in a number of characteristics of their skeleton, they were still on a much lower evolutionary level, comparable perhaps to the living tree-shrews (1). Modern primates are unique among mammals in their adaptation to life in the trees. Their capabilities of grasping and leaping allow rapid locomotion in this environment, which is in turn related to the large brain size they have developed. As far as we know, plesiadapiforms also spent most of their time in the trees. However, they lack adaptations for fast leaping as we see them in modern primates and were not capable of moving as quickly through the trees. In addition, their brain was still very small in comparison to modern primates. On the other hand, plesiadapiforms soon acquired traits that are unusual for later primates, especially enlarged incisors that are superficially similar to those of rodents. This suggests that the plesiadapiforms were not the direct ancestors of modern primates, but rather a branch that split off from the mainline of primate evolution (from today's point of view) at an early date. The picture is complicated by other still existing orders of mammals that must be close to primates on the evolutionary tree: The tree-shrews (order Scandentia), the colugos or "flying lemurs" (order Dermoptera), and perhaps the bats (order Chiroptera), although the latter are only distantly related according to recent molecular studies (2, 3). Where plesiadapiforms fit in between all of these is still hotly debated (4). However, many scientists are today reluctant to call these archaic forms primates and regard them instead as members of a separate order Plesiadapiformes (5).
Figure 1: Hypothetical restoration of the Early Paleocene Purgatorius, one of the earliest known primate-like mammals. From (28).
The most primitive known plesiadapiform, and one of the earliest, is the famous Purgatorius, an animal comparable to a small rat in size (6). Only the remains of the teeth and jaws are known for Purgatorius. Their structure is not far from the pattern that we would expect in the common ancestor of all placental mammals, presumably a small generalized insectivore. Yet the teeth of Purgatorius show some traits that are characteristic of both plesiadapiforms and true primates. Their design suggests an animal that fed not only on insects but also on some kind of soft plant material, probably fruits. Restorations often show Purgatorius scurrying under the feet of Late Cretaceous dinosaurs like Tyrannosaurus and Triceratops. Actually one species, Purgatorius ceratops, was named with this picture in mind. However, nearly all fossils of Purgatorius are from the Early Paleocene of North America. Only a single tooth has ever been found together with the remains of dinosaurs, and this tooth could be a contamination from Paleocene rock that was mixed by mistake with Cretaceous material when the collectors sieved the sediments. Even if no mixing happened during collecting, it is possible that something similar occurred nearly 65 million years ago: The deposits producing the single Purgatorius tooth were probably laid down during early Paleocene time by a river cutting deeply into Cretaceous sediments, mixing the fresh remains of Paleocene mammals with older fossils such as dinosaur teeth (7). Although unknown primate-like animals may have existed before the end of the Mesozoic, we still have no convincing evidence of them.
 If we want to
know more about early primate-like beings than jaws and teeth can
tell us, we have to take a look at later, more specialized forms.
The best known plesiadapiforms are the Plesiadapidae, one of the
most successful families of Paleocene mammals (1,
8). Plesiadapids were chipmunk- to
marmot-sized animals that had a superficially rodent-like
dentition, with a pair of robust, enlarged incisors that is
followed by a long gap without teeth (diastema) in later forms.
Whereas the lower incisors are relatively simply built and form a
kind of scoop together, the upper ones are more complicated, with
three separate cusps in front and another one behind, a
configuration that has been compared to mittens. Unlike the
enlarged incisors of rodents, the front teeth of plesiadapids
neither have a self-sharpening cutting edge nor are they
ever-growing, so they were probably utilized for one or several
purposes different from gnawing. In advanced plesiadapids the
cheek teeth are becoming flattened and the enamel is increasingly
crenulated, which suggests that these animals mainly subsisted on
a vegetable diet, perhaps on leafs and some fruits.
If we want to
know more about early primate-like beings than jaws and teeth can
tell us, we have to take a look at later, more specialized forms.
The best known plesiadapiforms are the Plesiadapidae, one of the
most successful families of Paleocene mammals (1,
8). Plesiadapids were chipmunk- to
marmot-sized animals that had a superficially rodent-like
dentition, with a pair of robust, enlarged incisors that is
followed by a long gap without teeth (diastema) in later forms.
Whereas the lower incisors are relatively simply built and form a
kind of scoop together, the upper ones are more complicated, with
three separate cusps in front and another one behind, a
configuration that has been compared to mittens. Unlike the
enlarged incisors of rodents, the front teeth of plesiadapids
neither have a self-sharpening cutting edge nor are they
ever-growing, so they were probably utilized for one or several
purposes different from gnawing. In advanced plesiadapids the
cheek teeth are becoming flattened and the enamel is increasingly
crenulated, which suggests that these animals mainly subsisted on
a vegetable diet, perhaps on leafs and some fruits.
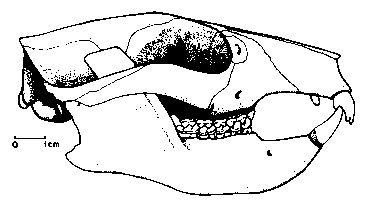
Figure 2: Skull of Plesiadapis tricuspidens from the Late Paleocene of France, vaguely resembling that of a rodent with its enlarged incisors and the long diastema in front of the cheek teeth. Note that the orbits of plesiadapiforms are not yet completely enclosed by bone, unlike those of true primates. From (8).
Skull and skeleton of plesiadapids are most completely known in the late Paleocene Plesiadapis. This genus probably arose in North America and colonized Europe on a landbridge via Greenland. Thanks to the abundance of the genus and to its rapid evolution, species of Plesiadapis play an important role in the zonation of Late Paleocene continental sediments and in the correlation of faunas on both sides of the Atlantic. Two remarkable skeletons of Plesiadapis, one of them nearly complete, have been found in lake deposits at Menat, France (8, 9, 10). Although the preservation of the hard parts is poor, these skeletons still show remains of skin and hair as a carbonaceous film - something unique among Paleocene mammals. Details of the bones are better preserved in fossils from Cernay, also in France, where Plesiadapis is one of the most common mammals. The rodent-like skull of Plesiadapis is relatively broad and flat, with a long snout and orbits still directed to the side, unlike the forward facing eyes of modern primates that enable three-dimensional vision (1, 6, 11). Although its braincase was small according to today's standards, it was larger than in the contemporary hoofed mammals, for instance. Plesiadapis had mobile limbs that terminated in strongly curved claws, and it sported a long bushy tail which is beautifully preserved in the Menat skeletons. The way of life of Plesiadapis has been much debated in the past. Climbing habits could be expected in a relative of the primates, but tree-dwelling animals are rarely found in such high numbers. Based on this and other evidence, some paleontologists have concluded that these animals were mainly living on the ground, like today's marmots and ground squirrels (8, 10). However, more recent investigations have confirmed that the skeleton of Plesiadapis is that of an adept climber, which can be best compared to tree squirrels or to tree-dwelling marsupials such as possums (1, 6).
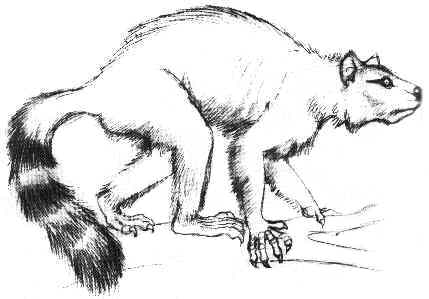
Figure 3: Restoration of the widespread genus Plesiadapis, showing the clawed hand and feet and the bushy tail. From (29).
It is noteworthy that early researchers surmised a close relationship between plesiadapids and the bizarre aye-aye of Madagascar, the only living primate with a rodent-like dentition. Later students have rejected such evolutionary ties, and the aye-aye is now regarded as a member of the same primate stock as lemurs and loris (12). Yet the aye-aye may be ecologically similar to a rare genus of plesiadapids, the squirrel-sized Chiromyoides from the Late Paleocene of Europe and North America (1, 6). Chiromyoides can be characterized as a super-robust version of Plesiadapis, with considerably shortened and deepened jaw and muzzle, and with extremely robust incisors that bear sharp cutting edges. Obviously its feeding mechanism had to withstand much higher forces than in other plesiadapids. Jaws and teeth of Chiromyoides closely resemble those of the aye-aye, which was sometimes called Chiromys in previous times - hence the name of the plesiadapid. The aye-aye uses its enlarged incisors to gnaw into wood in search of grubs. Chiromyoides may have occupied a similar niche in the forests of the Late Paleocene. This specialization would also explain its rarity in the mammal communities of that time. It may be of interest to note that a second group of Early Tertiary mammals, the Apatemyidae, evolved very similar adaptations, probably even approximating the aye-aye more closely than Chiromyoides.
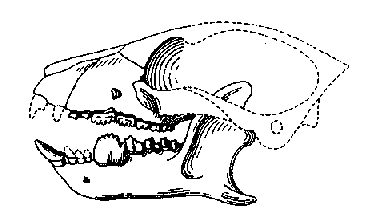 A second family of plesiadapiforms, the
Carpolestidae, is characteristic of Paleocene faunas in North
America, although it never becomes as dominant as the
plesiadapids (1, 6, 13, 14). Like the latter,
carpolestids have enlarged incisors, the lower ones simply built
and the upper ones mitten-shaped. However, they are much smaller,
ranging from the size of a mouse to that of a rat. Carpolestids
have developed their last lower premolars into enormous blades
with serrated cutting edges, a condition termed plagiaulacoid
that evolved independently in several groups of mammals, most
prominently in the archaic multituberculates.
In carpolestids these saw-like teeth worked against the also
enlarged but flattened and rasplike third and fourth premolar of
the upper jaw. This mechanism was probably used to process
vegetable diet with a high content of fibers such as fruits, nuts
or succulent shoots, a diet perhaps supplemented by some insects.
The evolution of this highly specialized adaptive complex can be
tracked in the successive North American genera Elphidotarsius,
Carpodaptes and Carpolestes that form the mainline
of carpolestid evolution in North America. The mouse-sized Elphidotarsius
from the Middle Paleocene of North America is still very similar
to primitive plesiadapids, but its last lower premolar already
foreshadows the blade-like form of advanced carpolestids. By Late
Paleocene time, the diagnostic traits of the family are well
established in Carpodaptes, and they are further refined
in Latest Paleocene species of Carpolestes ("fruit
stealer"), the last of the North American carpolestids.
A second family of plesiadapiforms, the
Carpolestidae, is characteristic of Paleocene faunas in North
America, although it never becomes as dominant as the
plesiadapids (1, 6, 13, 14). Like the latter,
carpolestids have enlarged incisors, the lower ones simply built
and the upper ones mitten-shaped. However, they are much smaller,
ranging from the size of a mouse to that of a rat. Carpolestids
have developed their last lower premolars into enormous blades
with serrated cutting edges, a condition termed plagiaulacoid
that evolved independently in several groups of mammals, most
prominently in the archaic multituberculates.
In carpolestids these saw-like teeth worked against the also
enlarged but flattened and rasplike third and fourth premolar of
the upper jaw. This mechanism was probably used to process
vegetable diet with a high content of fibers such as fruits, nuts
or succulent shoots, a diet perhaps supplemented by some insects.
The evolution of this highly specialized adaptive complex can be
tracked in the successive North American genera Elphidotarsius,
Carpodaptes and Carpolestes that form the mainline
of carpolestid evolution in North America. The mouse-sized Elphidotarsius
from the Middle Paleocene of North America is still very similar
to primitive plesiadapids, but its last lower premolar already
foreshadows the blade-like form of advanced carpolestids. By Late
Paleocene time, the diagnostic traits of the family are well
established in Carpodaptes, and they are further refined
in Latest Paleocene species of Carpolestes ("fruit
stealer"), the last of the North American carpolestids.
Figure 4: Skull of the Late Paleocene Carpodaptes. The saw-like premolar in the lower jaw characterizes the Carpolestidae. From (30).
 Figure 5: Restoration of the
Latest Paleocene Carpolestes simpsoni,
firmly grasping a branch with hands and feet. Credits D. M.
Boyer.
Figure 5: Restoration of the
Latest Paleocene Carpolestes simpsoni,
firmly grasping a branch with hands and feet. Credits D. M.
Boyer.
Carpolestids have long been known from little more than teeth and jaws, but a recently discovered nearly complete skeleton of Carpolestes simpsoni now sheds more light on their appearance and biology (15, 16). Carpolestes simpsoni was a small creature with a body weight of only 100 g, and obviously it was highly adapted to life in the trees. Its big toe could be opposed to the other toes, enabling the animal to firmly grasp smaller branches with its feet just like it could with its hands. The big toe of Carpolestes simpsoni carried a nail, something that allowed to develop far better tactile abilities than if the tip of the toe was covered by a claw. Nails were previously only known in true primates. Their presence in a plesiadapiform documents emphatically how these archaic forms were adapting to their arboreal environment, sometimes in ways also pursued by later primates. Unlike the first true primates, however, Carpolestes shows no adaptations for leaping.
Another family of highly specialized plesiadapiforms are the Picrodontidae, tiny animals such as the type genus Picrodus that occur with only a few species in the Middle and Late Paleocene of North America (1, 6). Picrodontids have long knifelike lower incisors, similar to those of other plesiadapiforms, but their cheek teeth are highly modified. The first upper and lower molars are extremely enlarged, and all molars form wide, shallow basins with heavily wrinkled enamel. Emphasis was clearly on increasing surface area in order to mash some kind of soft food. More abrasive food would have obliterated the low crowns of the teeth even during the very short life span that is typical of such small mammals. The specialized molars are so similar to those of certain recent bats that early researchers even considered picrodontids as possible members of that order, otherwise first known from the Early Eocene (17). Today this resemblance is regarded as convergent, but it provides important clues about the feeding habits of picrodontids. Like those bats, picrodontids probably fed on nectar and pollen, a highly nutritious diet that is very easy on the teeth, and perhaps also on tree sap and juicy fruit. A skull fragment of the picrodontid Zanycteris displays a relatively narrow muzzle. Any other adaptation of picrodontids to their particular ecological niche remains open to speculation.
Even smaller plesiadapiforms belong to the Micromomyidae (1, 6, 18). The family includes diminutive forms from the Late Paleocene to Early Eocene of the Rocky Mountain region which carry such appropriate names as Micromomys and Tinimomys (the suffix -mys meaning "mouse"). These animals may just have weighed about 30 g - distinctly less than the smallest living primate, the mouse-lemur Microcebus of Madagascar. An amazingly complete skeleton of a still unnamed micromomyid has recently been discovered in Latest Paleocene sediments. It suggests a tiny creature of only 20 g body weight that was highly adapted for climbing and may have been able to suspend itself from trees (15). Mammals of such small body weight are typically insectivorous, and in fact the sharp teeth of Micromomys confirm that this was a committed insect hunter.
Other plesiadapiforms seem to have harvested the diverse fruits of the widespread tropical to subtropical forests. Members of the Paromomyidae are characterized by molars that are squared off and flattened, forming large basins that would have been well suited for mashing fruit. Tree sap and insects may have complemented this diet (1, 6, 15, 19). The dental trends of paromomyids can be traced back to Paromomys from the Middle Paleocene of North America. In advanced forms such as Ignacius and Phenacolemur the central incisors become extremely enlarged and slender, and a large diastema develops behind them, like in plesiadapids. Both Ignacius and Phenacolemur were long-lived genera that range from the Late Paleocene to the Middle Eocene. Thus they were among the last surviving plesiadapiforms - living fossils in a world that was since long inhabited by true primates.
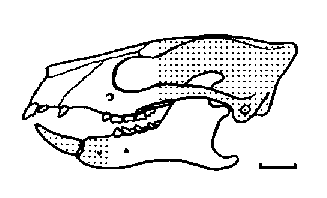
Figure 6: Skull of Ignacius graybullianus from the Latest Paleocene to Early Eocene of North America (stippled parts reconstructed). Note general resemblance of the dentition to Plesiadapis (Figure 2) but even longer incisors in the lower jaw. Scale bar 1 cm. From (11).
Paromomyids play an important role in a hypothesis from the 1990s which assumes close relationships between the plesiadapiforms and the colugos or "flying lemurs". The latter form one of the smallest existing orders of mammals, the Dermoptera, with just two species from Southeast Asia (1). Colugos have a gliding membrane that runs from behind their ears to the tip of their tail and that allows them to glide over large distances. The gliding membrane of colugos even extends between the digits of hand and foot, which enables them to control flight better than other gliding mammals. Because of similar hand structures in dermopterans and paromomyids, it has been claimed that the latter also possessed a gliding membrane. This and other evidence has led some researchers to the conclusion that plesiadapiforms are ancient dermopterans, today's colugos being the relics of a once diverse order of mammals that forms the sister group of primates (19, 20, 21). More complete fossils, however, do not confirm the idea that paromomyids were gliders. Instead paromomyids appear to have been squirrel-like creatures that could scamper both in the trees and on the ground (15, 22).
Even if no special relationships may exist between plesiadapiforms and colugos, other Paleocene mammals may prove to be early dermopterans. Among these are the plagiomenids, an enigmatic group that first appears in the Middle Paleocene of North America (1, 23). Plagiomenids are rare in most faunas, but they can dominate at individual localities, which may indicate particular habitat requirements. Fossils of the Latest Paleocene plagiomenid Planetetherium, for example, occur in abundance at a coal mine in Montana but are hardly known elsewhere. The animals must have found ideal conditions in the heavy forested or swampy environment represented by the mine deposits (24, 25). Interestingly, during Eocene times some plagiomenids even thrived on Ellesmere Island, the northernmost part of Canada that lies well within the Arctic Circle. Although climatic conditions where then of course much warmer than today, the animals living there must still have faced the winter darkness of such high latitudes (at least 76° North at that time) (1).
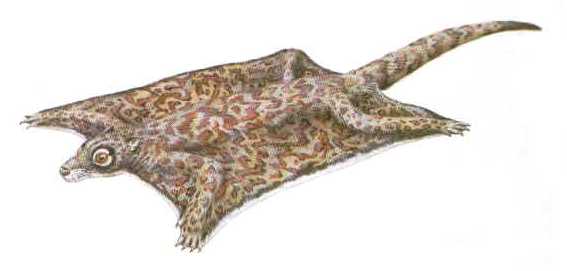
Figure 7: Hypothetical restoration of the Latest Paleocene Planetetherium as a glider similar to today's colugos or "flying lemurs". From (28).
The supposed link between plagiomenids and colugos is exclusively based on similarities of their dentitions, which have long been the only plagiomenid fossils. The lower incisors of plagiomenids, for instance, consist of two lobes, perhaps an early stage in the evolution towards the specialized comblike lower incisors that today's colugos use for grooming. Similarities of the cheek teeth, though somewhat vague, also suggest that plagiomenids may be early dermopterans. If this is the case, it would not be surprising to find gliding adaptations in Paleocene plagiomenids, given the fact that other mammals had evolved into full-fledged bats by the Early Eocene. This idea cannot be directly tested today with the available fossils. The link between plagiomenids and colugos has been weakened, however, by a recently discovered skull of the Latest Paleocene to Early Eocene Plagiomene, which shows some important anatomical differences in comparison to dermopterans (23). In face of this conflicting evidence, complete plagiomenid skeletons are urgently needed to solve the mystery of these intriguing animals.
Plesiadapiforms, and perhaps plagiomenids, belong to a first wave of primate-like mammals that flourished in the Paleocene. These archaic forms were replaced in the Eocene by true primates, which are called euprimates in technical jargon. The earliest known euprimates belong to the families Adapidae and Omomyidae (4). Both groups already possessed the advanced grasping and leaping adaptations and the correlated large brain that characterize modern primates. The adapids were similar and perhaps related to the lemurs of Madagascar and to the loris of tropical Africa and Asia. They mainly include larger animals that were presumably active during the day. Typical omomyids were small creatures with large eyes that suggest activity during the night. Omomyids may include the ancestors of the Southeast Asian tarsiers, nocturnal hunters of small prey that are great leapers and habitually cling to branches in a vertical position. Adapids and omomyids appear all of a sudden in the fossil record of Europe and North America at the beginning of the Eocene, which suggests they immigrated at that time from another part of the world, either from Africa or from Asia. Supposed euprimates have in fact been described from the Paleocene of these continents. Petrolemur and Decoredon from the Paleocene of China have been advanced as early members of the Adapidae and Omomyidae respectively. Both are poorly known, however, and it is not even sure that these animals are related to the primates at all (26). A more promising candidate was Altiatlasius from the Late Paleocene of Marocco, one of the few mammals that are known from the Paleocene of Africa. Originally described as an omomyid, Altiatlasius seemed to support the role of Africa as the center of primate evolution. Yet a recent study came to the result that Altiatlasius is no euprimate but belongs to a mainly European group of plesiadapiforms which dispersed into Africa across the ancient Tethys seaway (5, 27).
We thus still lack conclusive evidence about the early history of true primates - a history which began without question during Paleocene times, if not earlier (4). Nevertheless, we have ample evidence of the extraordinary success which their relatives, the plesiadapiforms, experienced in early Cenozoic faunas. As we have seen, the teeth of plesiadapiforms document a variety of feeding strategies, ranging from insectivores to generalized vegetarians and specialized pollen- and nectar-feeders, and we are just beginning to appreciate other aspects of their biology like locomotion. Hopefully, new discoveries will one day fill in the gap that separates these archaic forms from true members of our own order, the Primates.
First upload 14.04.02. Special thanks to J. Bloch and D. Boyer for supplying information on new plesiadapiform skeletons and for providing the Carpolestes restoration.
References: